
Di dalam sel, DNA berikatan dengan protein. Setiap molekul DNA dan protein terkaitnya disebut kromosom. Kromosom adalah bentuk kompak dari DNA. Pengemasan DNA ke dalam kromosom berfungsi untuk melindungi DNA dari kerusakan. Hanya DNA yang dikemas ke dalam kromosom yang dapat ditransmisikan secara efisien ke kedua sel anak ketika sel membelah. Akhirnya, kromosom bisa dibilang sebagai suatu organisasi besar untuk setiap molekul DNA.

Dalam sel eukariotik, daerah tertentu dari DNA dengan protein terkait disebut kromatin. Sebagian besar protein terkait ini merupakan protein kecil yang disebut histon. Protein non-histon, juga terkait dengan kromosom eukariotik. Protein ini mencakup banyak protein pengikat DNA yang mengatur replikasi, perbaikan, rekombinasi, dan transkripsi DNA seluler.
Komponen protein kromatin melakukan fungsi penting lainnya: memadatkan DNA. Kebanyakan pemadatan dalam sel manusia (dan semua sel eukariotik lainnya) adalah hasil dari ikatan DNA dengan histon untuk membentuk struktur yang disebut nukleosom.

Urutan genom dan keanekaragaman kromosom
Kromosom dapat berbentuk sirkuler (lingkaran) atau linier (garis lurus). Meskipun sebagian besar sel prokariotik memiliki satu kromosom sirkuler saja, sekarang diketahui ada banyak contoh sel prokariotik yang memiliki banyak kromosom, punya kromosom linier, atau bahkan keduanya. Sebaliknya, semua sel eukariotik memiliki banyak kromosom linier.
Setiap sel punya karakter kromosom tersendiri. Sel prokariotik biasanya hanya memiliki satu salinan lengkap dari kromosom yang dikemas ke dalam struktur yang disebut nukleoid. Prokariota juga sering membawa satu atau lebih DNA sirkuler independen yang lebih kecil, yang disebut plasmid.
Mayoritas sel eukariotik adalah diploid; mengandung dua salinan dari setiap kromosom. Dua salinan dari kromosom tertentu disebut homolog—satu diturunkan dari masing-masing orang tua. Tetapi tidak semua sel dalam organisme eukariotik adalah diploid; sebagian sel eukariotik adalah haploid atau poliploid. Sel haploid mengandung satu salinan dari setiap kromosom dan terlibat dalam reproduksi seksual (misalnya, sperma dan sel telur adalah sel haploid). Sel poliploid memiliki lebih dari dua salinan dari setiap kromosom (misalnya, megakariosit). Berapa pun jumlahnya, kromosom eukariotik selalu terkandung dalam organel yang terikat membran yang disebut nukleus.

Ukuran genom (panjang DNA yang terkait dengan satu komplemen kromosom haploid) bervariasi antara organisme yang berbeda. Sel prokariotik biasanya memiliki genom <10 Mb (1 megabase/Mb = 1.000.000 base pair/bp). Genom eukariota sel tunggal biasanya <50 Mb. Organisme multiseluler bahkan memiliki genom yang lebih besar yang dapat mencapai ukuran> 100.000 Mb.
Kepadatan gen adalah jumlah rata-rata gen per megabase DNA genom. Organisme yang lebih kompleks mengalami penurunan kepadatan gen. Ada dua faktor yang berkontribusi pada penurunan kepadatan gen yang diamati pada sel eukariotik: (1) peningkatan ukuran gen dan (2) peningkatan DNA antar gen, yang disebut sekuens intergenik.

Gen penyandi protein pada eukariota sering memiliki daerah penyandi protein yang terputus-putus karena diselingi oleh daerah yang tidak menyandi protein yang disebut intron. Nantinya, intron akan dikeluarkan dari RNA setelah transkripsi dalam proses yang disebut penyambungan/splicing RNA.

Ada dua jenis sekuens intergenik: unik dan berulang. Penyusun utama sekuens unik adalah sekuens DNA yang diperlukan untuk mengarahkan dan mengatur transkripsi, yang disebut sekuens regulator. Sekuens unik pada mansuia juga mencakup banyak sekuens yang nonfungsional, meliputi gen mutan, fragmen gen, dan pseudogen. Gen mutan dan fragmen gen muncul dari mutagenesis acak sederhana atau kesalahan dalam rekombinasi DNA. Pseudogen muncul dari aksi enzim yang disebut reverse transcriptase yang menyalin RNA menjadi DNA untai ganda.
Sementara itu sekuens DNA berulang ada dua: DNA mikrosatelit dan genom-wide repeats. DNA mikrosatelit terdiri dari urutan yang sangat pendek (<13 bp), berulang secara tandem. Genom wide-repeats jauh lebih besar daripada DNA mikrosatelitnya. Setiap unit genom wide-repeats memiliki panjang >100 bp dan banyak yang >1 kb. Fitur umum mereka adalah bahwa semua adalah bentuk elemen transposabel yang dapat “berpindah” dalam genom dari satu tempat ke tempat lain.

Duplikasi dan segregasi kromosom
Kromosom eukariotik membutuhkan sentromer, telomer, dan origin of replication untuk dipertahankan selama pembelahan sel. Origin of replication mengarahkan duplikasi DNA kromosom. Sentromer bertindak sebagai “pegangan” untuk pergerakan kromosom hasil duplikasi ke dalam sel anak. Sementara telomer melindungi dan menggandakan ujung kromosom linier.

Origin of replication adalah daerah di mana perangkat replikasi DNA berkumpul dan replikasi dimulai. Sentromer diperlukan untuk pemisahan kromosom setelah replikasi DNA. Dua salinan dari setiap kromosom yang direplikasi disebut sister-kromosom, dan selama pembelahan sel mereka harus dipisahkan dengan satu salinan pergi ke masing-masing dari dua sel anak. Seperti origin of replication, sentromer mengarahkan pembentukan kompleks protein rumit yang disebut kinetokor. Kinetokor berkumpul di setiap DNA sentromer, dan sebelum pemisahan kromosom, kinetokor mengikat filamen protein yang disebut mikrotubulus yang akhirnya menarik sister-kromosom menjauh dari satu sama lain dan ke dalam dua sel anak. Telomer terletak di kedua ujung kromosom linier. Telomer memfasilitasi replikasi akhir melalui perekrutan DNA polimerase yang tidak biasa yang disebut telomerase.


Peristiwa yang diperlukan untuk satu putaran pembelahan sel secara kolektif dikenal sebagai siklus sel. Sebagian besar pembelahan sel eukariotik mempertahankan jumlah kromosom dalam sel anak yang ada dalam sel induk. Jenis pembelahan ini disebut pembelahan sel mitosis.

Siklus sel mitosis dapat dibagi menjadi empat fase: G1, S, G2, dan M.
Replikasi kromosom terjadi selama sintesis, atau fase S, dari siklus sel, menghasilkan duplikasi setiap kromosom. Setiap kromosom dari pasangan yang digandakan disebut kromatid, dan dua kromatid dari pasangan tertentu disebut sister-kromatid. Sister-kromatid disatukan setelah duplikasi melalui proses yang disebut kohesi sister-kromatid, dan keadaan tertambat ini dipertahankan sampai kromosom berpisah satu sama lain. Kohesi sister-kromatid dimediasi oleh protein yang disebut kohesin.
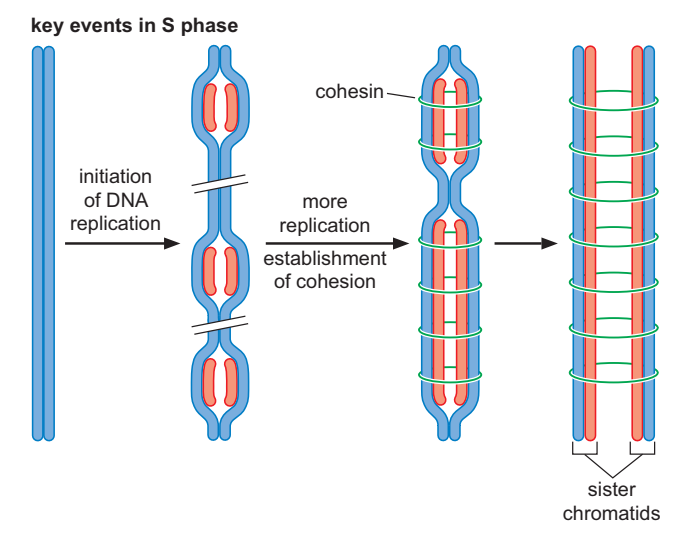
Segregasi kromosom terjadi selama mitosis, atau fase M, dari siklus sel. Tiga peristiwa besar terjadi selama mitosis: (1) Setiap pasangan sister-kromatid terikat pada suatu struktur yang disebut mitotic spindle. Struktur ini terdiri dari serat protein panjang yang disebut mikrotubulus yang melekat pada salah satu dari dua pusat pengorganisasian mikrotubulus (juga disebut sentrosom). Perlekatan kromatid ke mikrotubulus dimediasi oleh kinetokor yang dipasang pada setiap sentromer. (2) Kohesi antara kromatid dirombak melalui proteolisis. Setelah kohesi dihilangkan, (3) sister-kromatid dipisahkan ke kutub berlawanan dari mitotic spindle.

Ada dua keadaan utama untuk kromosom: Interfase dan fase M. Kromosom terkondensasi secara maksimal dalam fase M dan terdekondensasi sepanjang sisa siklus sel (G1, S, dan G2 dalam sel mitosis). Bersama-sama, tahap dekondensasi ini disebut sebagai interfase. Protein SMC (structural maintenance of chromosome) memediasi kohesi sister-kromatid dan kondensasi kromosom. Kondensasi kromosom yang menyertai segregasi kromosom juga memerlukan kompleks yang mengandung SMC yang disebut kondensin.

Mitosis terjadi dalam beberapa tahap:
(1) Profase: aksi kondensin dan topoisomerase tipe II (untuk membantu menguraikan kromosom) mendorong kromosom untuk memadat menjadi bentuk yang sangat kompak yang diperlukan untuk segregasi. Pada akhir profase, selubung inti pecah.
(2) Metafase: mitotic spindle terbentuk dan kinetokor sirter-kromatid menempel pada mikrotubulus.
(3) Anafase: hilangnya kohesi sister-kromatid sehingga terjadi pemisahan.
(4) Telofase: hilangnya kondensasi kromosom dan pembentukan kembali membran nukleus di sekitar dua populasi kromosom yang terpisah.
(5) Sitokinesis: peristiwa akhir dari siklus sel di mana membran sel yang mengelilingi dua inti menyempit dan akhirnya benar-benar terpisah menjadi dua sel anak. Semua molekul DNA adalah untai ganda.

Dua fase yang tersisa dari siklus sel mitosis adalah fase gap. G1 terjadi sebelum sintesis DNA, dan G2 terjadi antara fase S dan fase M. Fase gap dari siklus sel menyediakan waktu bagi sel untuk mencapai dua tujuan: (1) untuk mempersiapkan fase berikutnya dari siklus sel dan (2) untuk memeriksa bahwa fase sebelumnya dari siklus sel telah diselesaikan dengan tepat. Jika ada masalah dengan langkah sebelumnya dalam siklus sel, pos pemeriksaan siklus sel menghentikan siklus sel untuk menyediakan waktu bagi sel untuk menyelesaikan langkah itu.
Siklus sel meiosis meliputi fase G1, S, dan fase G2 memanjang. Sel yang masuk meiosis harus diploid. Setelah replikasi DNA, sister-kromatid homolog berpasangan satu sama lain untuk membentuk struktur dengan empat kromosom terkait. Selama berpasangan, kromatid dari homolog yang berbeda bergabung kembali untuk membentuk hubungan antara kromosom homolog yang disebut kiasma. Pada metafase I dua kinetokor dari setiap pasangan sister-kromatid menempel pada satu kutub spindel mikrotubulus. Kinetokor kromatid homolog menempel pada kutub yang berlawanan, menciptakan tegangan yang ditahan oleh kiasma antara homolog dan kohesi antara lengan seister-kromatid. Anafase I ditandai dengan hilangnya kohesi sister-kromatid di sepanjang lengan kromosom sehingga memungkinkan homolog rekombinasi terpisah satu sama lain. Sister-kromatid tetap melekat melalui kohesi di sentromer. Meiosis II sangat mirip dengan mitosis. Selama metafase II meiosis, dua spindel meiosis terbentuk. Seperti pada metafase mitosis, kinetokor yang terkait dengan setiap pasangan sister-kromatid menempel pada kutub yang berlawanan dari spindel meiosis. Selama anafase II, kohesi sentromer yang tersisa antara sister-kromatid hilang, dan sister-kromatid terpisah satu sama lain. Empat set kromosom yang terpisah kemudian dikemas menjadi inti dan dipisahkan menjadi empat sel untuk membuat empat spora atau gamet yang haploid.

Nukleosom
Nukleosom adalah blok bangunan kromosom. Setiap nukleosom terdiri dari 8 protein histon inti (histone core) dan DNA yang melilit mereka. DNA antara setiap nukleosom disebut DNA penghubung (linker DNA). DNA yang paling erat ikatannya dengan nukleosom disebut DNA inti (core DNA).

Sel eukariotik umumnya mengandung lima histon yang melimpah: H1, H2A, H2B, H3, dan H4. Histon H2A, H2B, H3, dan H4 adalah histon inti, dan dua salinan dari masing-masing histon ini membentuk inti protein yang membungkus DNA nukleosom. Histon H1 bukan bagian dari partikel inti nukleosom. Sebaliknya, ia mengikat DNA penghubung dan disebut sebagai histon penghubung. Histon inti juga merupakan protein yang relatif kecil dengan ukuran mulai dari 11 hingga 15 kilodalton (kDa). Histon H1 sedikit lebih besar dengan ukuran 21 kDa. Pada histon inti terdapat regio yang conserved, disebut dengan domain lipatan histon (histone-fold domain). Selain itu masing-masing histon inti memiliki ujung amino N terminal, yang disebut ekor (N-terminal tail). Ekor adalah tempat modifikasi pascatranslasi ekstensif yang dapat mengubah fungsi nukleosom suatu individu.

Perakitan nukleosom dimulai dengan pembentukan tetramer H32.H42. Tetramer kemudian berikatan dengan DNA untai ganda. Tetramer H32.H42 yang terikat pada DNA merekrut dua salinan dimer H2A.H2B untuk menyelesaikan perakitan nukleosom.

Nukleosom memiliki perkiraan sumbu simetri rangkap dua, yang disebut diad axis. Hal ini dapat divisualisasikan dengan memikirkan wajah piringan oktamer sebagai jam, dimana titik tengah DNA 147 bp terletak pada posisi jam 12, sementara ujung DNA diproyeksikan pada arah jam 11 dan jam 1. Sebuah garis yang ditarik dari jam 12 sampai jam 6 melalui tengah piringan adalah diad axis. Rotasi nukleosom sebesar 180 ° di sumbu ini akan memperlihatkan tampilan nukleosom yang hampir identik dengan yang diamati sebelum rotasi.

Asosiasi DNA dengan nukleosom dimediasi oleh sejumlah besar (sekitar 40) ikatan hidrogen antara histon dan DNA. Mayoritas ikatan hidrogen ini berada di antara protein dan atom oksigen di tulang punggung fosfodiester dekat alur kecil DNA.
Ekor histon muncul dari inti nukleosom pada posisi tertentu. Ekor H2B dan H3 muncul pada jarak yang kira-kira sama satu sama lain di sekitar cakram oktamer (pada jam 1 dan 11 untuk ekor H3 dan jam 4 dan 8 untuk H2B. Ekor H2A muncul pada jam 5 dan 7. dan ekor H4 pada jam 3 dan 9.

Struktur kromatin tingkat tinggi
Studi awal tentang kromosom membagi regio kromosom menjadi dua kategori: eukromatin dan heterokromatin. Heterokromatin dicirikan dengan pewarnaan yang padat dengan berbagai zat warna dan kenampakan yang lebih kental, sedangkan eukromatin memiliki karakteristik yang berlawanan, pewarnaan yang buruk dengan zat warna dan memiliki struktur yang relatif terbuka. Daerah heterokromatik kromosom memiliki ekspresi gen yang sangat terbatas. Sebaliknya, daerah eukromatik menunjukkan tingkat ekspresi gen yang lebih tinggi.
Setelah nukleosom terbentuk, langkah selanjutnya dalam pengemasan DNA adalah pengikatan histon H1. Histon H1 mengikat dua heliks DNA. Histon H1 mengikat DNA penghubung di salah satu ujung nukleosom dan heliks DNA inti (bagian tengah 147 bp yang diikat oleh inti oktamer histone). Pengikatan H1 menstabilkan struktur kromatin tingkat tinggi. Histone H1 menginduksi pembungkus DNA yang lebih ketat di sekitar nukleosom.


Penambahan histon H1 menghasilkan DNA nukleosom yang membentuk serat kromatin 30-nm. Ada dua model untuk struktur serat 30 nm.
(1) Model solenoida: DNA nukleosomal membentuk superhelix yang mengandung kira-kira enam nukleosom per putaran. Perhatikan bahwa DNA penghubung tidak melewati sumbu pusat superhelix dan bahwa sisi serta titik masuk dan keluar nukleosom relatif tidak dapat diakses.
(2) Model “zigzag”: DNA penghubung melewati sumbu tengah serat, dan sisi-sisi serta titik masuk dan keluar lebih mudah diakses.

Ekor terminal amino histone diperlukan untuk membentuk dan menstabilkan serat 30-nm dengan berinteraksi dengan nukleosom yang berdekatan.

Sebagian besar DNA dikemas ke dalam loop besar serat 30-nm yang ditambatkan ke bagian inti di dasarnya. Bagian DNA yang aktif (misalnya, situs transkripsi atau replikasi DNA) biasanya terletak pada serat 10-nm atau DNA telanjang (naked DNA).

Pengaturan struktur kromatin
Nukleosom dapat dipindahkan, atau cengkeramannya pada DNA dapat dilonggarkan untuk memungkinkan protein lain mengakses DNA. Ikatan antara histon oktamer dengan DNA-nya bersifat dinamis. Stabilitas interaksi histone oktamer-DNA dipengaruhi oleh kompleks protein besar yang disebut kompleks remodeling nukleosom.
Ada tiga tipe dasar perubahan nukleosom: (1) Pergerakan nukleosom dengan menggeser (sliding) di sepanjang molekul DNA akan mengekspos bagian untuk protein pengikat DNA. (2) Kompleks remodeling nukleosom juga dapat mengeluarkan (ejecting) nukleosom dari DNA yang menciptakan daerah DNA bebas nukleosom yang lebih besar. (3) Sebuah subset kompleks remodeling nukleosom mengkatalisis pertukaran (exchange) dimer H2A/H2B dengan dimer modifikasi atau varian H2A/H2B (misalnya, H2A-X).

Perakitan nukleosom
Nukleosom dirakit segera setelah replikasi DNA. Langkah pertama dalam perakitan nukleosom adalah pengikatan tetramer H3.H4 ke DNA. Setelah tetramer terikat, dua dimer H2A.H2B bergabung untuk membentuk nukleosom akhir. H1 bergabung dengan kompleks ini selama pembentukan rakitan kromatin tingkat tinggi. Histon lama akan tetap hadir pada kedua kromosom anak.

Perakitan nukleosom membutuhkan histon “pendamping”. Setelah garpu replikasi (replication fork) berlalu, faktor perakitan kromatin “mendampingi” tetramer H3.H4 bebas (misalnya, CAF-I) dan dimer H2A.H2B (NAP-I) ke situs DNA yang baru direplikasi. Begitu berada di DNA yang baru direplikasi, faktor-faktor ini mentransfer histon yang didampinginya ke DNA.

Kesimpulan
Genom suatu makhluk hidup disimpan dengan aman dalam bentuk kromosom. Kromosom disusun oleh kumpulan kromatin yang menggulung-gulung secara rapi, dan kromatin dibangun oleh susunan nukleosom yang berderet-deret. Di dalamnya berperan protein histon, baik inti maupun ekor. Selama fase interfase siklus sel, kromosom akan dijaga dalam bentuk lepas, mengalami pengecekan dan perbaikan jika ada kerusakan dan menggandakan diri melalui replikasi. Ketika masuk ke fase pembelahan, kromosom akan dipadatkan. Pembelahan mitosis akan menghasilkan dua sel yang diploid, persis sama dengan induknya, sementara pembelahan meiosis akan menghasilkan sel yang haploid, separoh kromosom induknya. Semua proses ini memastikan bahwa DNA makhluk hidup akan senantiasa terjaga dan melaksanakan fungsi sesuai fitrahnya.
Wallahu’alam.
Referensi: Watson et al. 2013. Molecular Biology of the Gene, 7th edition. New York: Cold Spring Harbor Laboratory Press.
Cover image: https://cdnb.artstation.com/
